
MHC对免疫应答中免疫细胞相互作用的限制(约束)作用
2011-11-27 10:12:13 来源: 作者: 评论:0 点击:
MHC另一个重要的生物学功能是约束免疫应答过程中各类免疫细胞的相互作用,又称为MHC的约束性(MHc restriction),包括免疫应答感应阶段Mφ-Th之间,反应阶段Th-B之间,以及效应阶段Tc-靶细胞之间的相互作用。MHc I类和Ⅱ类抗原分别对不同细胞起约束作用。
(一)Mφ、T、B细胞相互作用过程中的MHC约束性
Rosenthal和Shevach(1973)首先在豚鼠中观察到T细胞只能被具有相同MHc I区基因(Ⅱ类基因)的抗原提呈细胞所激活(表6-13)。同年Katz等人也发现Th与B细胞相互作用时,只有在两者MHc I区(Ⅱ类基因)相同的条件才会出现协作应(表6-14)。
表6-13 豚鼠T细胞对胸腺信赖抗原的免疫应答中Mφ提呈抗原与Ia抗原的关系
OVA预处理T细胞OVA加MφDNA合成的增加
品系2品系2++++
品系13品系13++++
品系2品系13—
品系13品系2—
(2*13)F1品系2+++
(2*13)F1品系13+++
(2*13)F1(2*13)F1++++
注:豚鼠用卵清蛋白(OVA)加完全弗氏佐剂(CFA)免疫,腹腔渗出物分离的T细胞即为OVA预处理T细胞,在体 外与OVA加Mφ-起培养,用3H-胸腺嘧啶核苷掺入法作为T细胞特异性免疫应答。品系2和13仅在MHC的I区不同
表6-14 小鼠T-B细胞协作中受I区基因限制
DNP-KLH抗原预致敏B细胞的品系BGG载体预致敏T细胞的品系受照射(a*b)F1受体对DNP-BGG抗体应答再次反应
aa++++
bb++++
ab-
ba-
b(a*b)F1++++
a(a*b)F1++++
(a*b)F1a++++
(a*b)F1b++++
注:(1)DNP-DLH:二硝基苯酚-钥孔墄血兰素;DGG:牛r球蛋白。
(2)DNP-KLH预致敏B细胞为DNP-KLH免疫小鼠的脾细胞经抗Thy-1加补体处理去除T细胞而获得。
(3)BGG免疫小鼠脾细胞除去B细胞后为BGG载体预致敏T细胞。
(4)将DNP-KLH预致敏B细胞和BGG预致敏T细胞输入经照射后(a*b)F1受体,再用DNP-BGG抗原刺激,检测机体对DNP抗体应答的再次反应。
(二)MHC对Tc杀伤病毒感染靶细胞的约束
1.Zinkernagel-Doherty现象 Zinkernagel和Doherty(1974)道德证明受牛痘病毒感染的CBA(H-2k)小鼠中的Tc只能杀伤H-2单体型相同的病毒感染靶细胞,而不能杀死牛痘苗病毒感的H-2b细胞,称为“Zinkernagel-Doherty phenomenon”。1975年Doherty用淋巴细胞脉络膜脑膜炎病毒(lymphocyte-choriomeningitis virus,LCM病毒)感染H-2d小鼠,取出Tc在体外只能杀伤LCM病毒感染的H-2d单体型细胞,不能杀伤LCM病毒感染的H-2k细胞(见图6-18)。上述实验表明,Tc对于只具有MHC抗原或病毒抗原中单独一种抗原的靶细胞都不起杀伤作用。

图6-18 MHC对Tc杀伤病毒感染靶细胞的约束作用
2. MHC I类抗原对Tc杀伤靶细胞的限制作用 参与免疫应答杀伤相(效应相)的H-2抗原是由K、D区决定的,I区并不参与,这不同于前述的免疫应答感应阶段中Mφ-Th,Th-B细胞之间相互作用受I区控制(表6-15)。
用三硝基苯(TNP)修饰自体的脾细胞为靶细胞也同样证实了MHC对Tc杀伤TNP修饰靶细胞的约束现象。在人类杀伤病毒感染或半抗原修饰的靶细胞也同样受到I类抗原的约束,如McMicheal等(1977)发现杀伤流感病毒感染的靶细胞主要受HLA-B位点抗原的约束,Dickmeiss(1977)实验表明杀伤DNFB致敏淋巴细胞诱导Tc杀伤靶细胞中,HLA-A抗原必须一致。
由于自身MHC约束Tc细胞杀伤靶细胞的特异性,使体内受病毒感染或癌肿恶变的靶细胞得以迅速有效地清除,从这个意义上来讲,MHC参与机体抗感染及免疫监视功能。
Tc对同种异体靶细胞的杀伤作用不受自身MHC的约束,Longo(1982)认为体内存在着两类不同的T应答细胞:一类针对外来抗原+自身MHC抗原发生免疫应答;另一类对同种异体细胞发生免疫应答。
表6-15 小鼠特异性Tc细胞的杀伤作用与靶细胞H-2的关系
靶细胞感染的病毒靶细胞H-2遗传背景杀伤作用
KID
LCMskd+
LCMqpq-
LCMsss+
LCMddd+
LCMkkk-
仙台病毒skd-
注:小鼠特异性Tc细胞取自受LCM病毒致敏、H-2型别为KsIkDd小鼠的脾脏
八十年代初,采用基因转染技术,将H-2b及H-2d的I类基因转染小鼠白血病细胞系(L细胞系)。这些受I类基因转染的细胞表面有I类抗原的表达。特异性Tc不能杀伤未经I类基因转染的病毒感染L细胞,但可杀伤经I类基因转染的病毒感染L细胞,表明转染后所表达的I类抗原起约束作用。Ozato等切割I类基因的不同片断进行基因杂交,并将其转染到L细胞系,成功地得到嵌合I类抗原分子,如α1及α2结构域取自与杀伤细胞I类基因相同的基因片段,而α3、穿膜及胞浆区取自与杀伤细胞I类基因不同基因片段,则杀伤细胞可杀伤病毒感染的靶细胞,所之则不然。上述试验证明,α1及α2结构域是参与杀伤和约束的有效部位。
利用突变品系小鼠研究发现靶细胞I类抗原突变分子的微小变化便能改变Tc细胞对它的识别。使如H-2kbml突变株(b为单倍体,m为突变)来自H-2Kb突变品系,只有个别氨基酸的差异,Kb约束病毒抗原特异性的CTL就不能识别和裂解多种病毒感染的Kb突变的bm1靶细胞,表明1~3个氨基酸的变化足以使I类抗原失去原有的约束作用(表6-16)。
3.MHC对Tc杀伤病毒感染靶细胞约束的机制 目前关于MHC对Tc细胞杀伤病毒感染靶细胞约束的机制一般认为是通过联合识别(associative recognition),即Tc表面一个受体识别MHC编码的抗原与病毒抗原的复合物。
Benacerraf认为MHC I类Ⅱ类抗耕牛具有双重功能,在诱导阶段通过其结合部位选择抗原决定基,激活Th、Ts、Tc等效应细胞;在效应阶段,靶细胞上的I类和Ⅱ类抗原又约束Th、Ts、Tc发挥效应。
(三)载体效应(cerrier effect)
Benacerraf认为载体不是单纯起运载半抗原的作用,还具有载体特异性。载体处于耐受状态的动物,虽给予半抗原载体复合物,也不能诱导产生抗半抗原的抗体。对半抗原再次应答的发生有赖于对半原记忆的B细胞和对载体记忆T细胞同时存在时才能发生(图6-19)。
表6-16 K(b)-约束的CTL(a)对Kb(b)突靶细胞上抗原的识别
CTL针对病毒抗原对Kb突变靶细胞的裂解
bm5(1)(c)bm6(2)bm9(3)bm3(2)bm8(3)bm11(1)bm1(3)
LCM病毒++++ +++ -
小鼠脱脚病病毒±+ + -
SV40++ ++++-
Moloney氏病毒++++++++-++-
Seudai病毒++++++++--
痘苗病毒++ ++ -
VSV++ (+)+ -
甲型流感病毒++ (+)- -
注:(a)CTL是Tc细胞,取自各种病毒致敏的小鼠脾,其本身的H-2型别为Kb;
(b)所有品系自B6突变而来,b为单倍型,m为突变;
(c)突变后氨基酸结构改变的数目
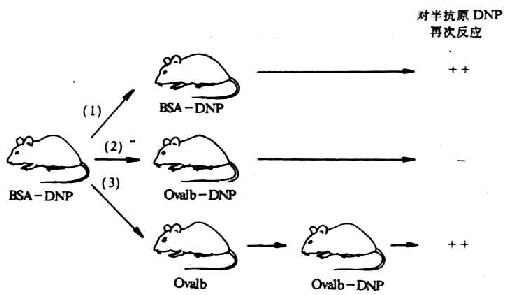
图6-19 载体效应
注:①第二次注射与第一次相同的半抗原(DNP)和载体(牛血清白蛋白BSA) ,结果产生对DNP的再次反应。
②第二次反注射的抗原中,半抗原与第一次相同,载体(卵白蛋白ovalbumin)与第一次注射抗原的载体不同,结果不产生再次反应;
③第一次注射后,单独注射卵白蛋白,经过一定时间后再注射载体为卵白蛋白、半抗原为DNP的抗原,结果产生再次反应。
Mitchison等载体效应的过继转移(adoptive transfer of carrier effect)试验,证实了上述关于载体效应的结论(图6-20)。

图6-20 载体效应的过继转移试验(Mitchison)
Raff等载体效应阻断实验进一步证实T细胞是载体特异的载体反应细胞(carrier-reactive cell),B细胞是半抗原反应细胞(hapten-reactive cell)(图6-21)。
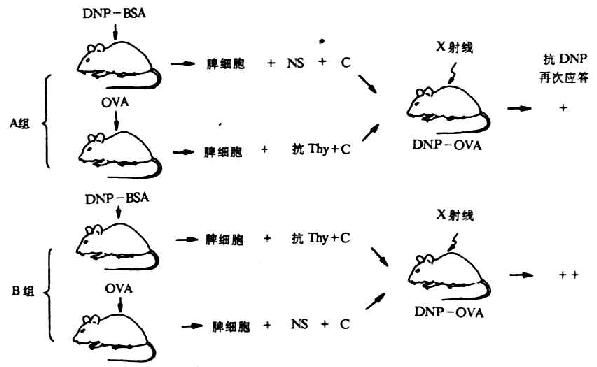
图6-21 半抗原、载体反应淋巴细胞的鉴定(Raff等)
(一)Mφ、T、B细胞相互作用过程中的MHC约束性
Rosenthal和Shevach(1973)首先在豚鼠中观察到T细胞只能被具有相同MHc I区基因(Ⅱ类基因)的抗原提呈细胞所激活(表6-13)。同年Katz等人也发现Th与B细胞相互作用时,只有在两者MHc I区(Ⅱ类基因)相同的条件才会出现协作应(表6-14)。
表6-13 豚鼠T细胞对胸腺信赖抗原的免疫应答中Mφ提呈抗原与Ia抗原的关系
OVA预处理T细胞OVA加MφDNA合成的增加
品系2品系2++++
品系13品系13++++
品系2品系13—
品系13品系2—
(2*13)F1品系2+++
(2*13)F1品系13+++
(2*13)F1(2*13)F1++++
注:豚鼠用卵清蛋白(OVA)加完全弗氏佐剂(CFA)免疫,腹腔渗出物分离的T细胞即为OVA预处理T细胞,在体 外与OVA加Mφ-起培养,用3H-胸腺嘧啶核苷掺入法作为T细胞特异性免疫应答。品系2和13仅在MHC的I区不同
表6-14 小鼠T-B细胞协作中受I区基因限制
DNP-KLH抗原预致敏B细胞的品系BGG载体预致敏T细胞的品系受照射(a*b)F1受体对DNP-BGG抗体应答再次反应
aa++++
bb++++
ab-
ba-
b(a*b)F1++++
a(a*b)F1++++
(a*b)F1a++++
(a*b)F1b++++
注:(1)DNP-DLH:二硝基苯酚-钥孔墄血兰素;DGG:牛r球蛋白。
(2)DNP-KLH预致敏B细胞为DNP-KLH免疫小鼠的脾细胞经抗Thy-1加补体处理去除T细胞而获得。
(3)BGG免疫小鼠脾细胞除去B细胞后为BGG载体预致敏T细胞。
(4)将DNP-KLH预致敏B细胞和BGG预致敏T细胞输入经照射后(a*b)F1受体,再用DNP-BGG抗原刺激,检测机体对DNP抗体应答的再次反应。
(二)MHC对Tc杀伤病毒感染靶细胞的约束
1.Zinkernagel-Doherty现象 Zinkernagel和Doherty(1974)道德证明受牛痘病毒感染的CBA(H-2k)小鼠中的Tc只能杀伤H-2单体型相同的病毒感染靶细胞,而不能杀死牛痘苗病毒感的H-2b细胞,称为“Zinkernagel-Doherty phenomenon”。1975年Doherty用淋巴细胞脉络膜脑膜炎病毒(lymphocyte-choriomeningitis virus,LCM病毒)感染H-2d小鼠,取出Tc在体外只能杀伤LCM病毒感染的H-2d单体型细胞,不能杀伤LCM病毒感染的H-2k细胞(见图6-18)。上述实验表明,Tc对于只具有MHC抗原或病毒抗原中单独一种抗原的靶细胞都不起杀伤作用。
图6-18 MHC对Tc杀伤病毒感染靶细胞的约束作用
2. MHC I类抗原对Tc杀伤靶细胞的限制作用 参与免疫应答杀伤相(效应相)的H-2抗原是由K、D区决定的,I区并不参与,这不同于前述的免疫应答感应阶段中Mφ-Th,Th-B细胞之间相互作用受I区控制(表6-15)。
用三硝基苯(TNP)修饰自体的脾细胞为靶细胞也同样证实了MHC对Tc杀伤TNP修饰靶细胞的约束现象。在人类杀伤病毒感染或半抗原修饰的靶细胞也同样受到I类抗原的约束,如McMicheal等(1977)发现杀伤流感病毒感染的靶细胞主要受HLA-B位点抗原的约束,Dickmeiss(1977)实验表明杀伤DNFB致敏淋巴细胞诱导Tc杀伤靶细胞中,HLA-A抗原必须一致。
由于自身MHC约束Tc细胞杀伤靶细胞的特异性,使体内受病毒感染或癌肿恶变的靶细胞得以迅速有效地清除,从这个意义上来讲,MHC参与机体抗感染及免疫监视功能。
Tc对同种异体靶细胞的杀伤作用不受自身MHC的约束,Longo(1982)认为体内存在着两类不同的T应答细胞:一类针对外来抗原+自身MHC抗原发生免疫应答;另一类对同种异体细胞发生免疫应答。
表6-15 小鼠特异性Tc细胞的杀伤作用与靶细胞H-2的关系
靶细胞感染的病毒靶细胞H-2遗传背景杀伤作用
KID
LCMskd+
LCMqpq-
LCMsss+
LCMddd+
LCMkkk-
仙台病毒skd-
注:小鼠特异性Tc细胞取自受LCM病毒致敏、H-2型别为KsIkDd小鼠的脾脏
八十年代初,采用基因转染技术,将H-2b及H-2d的I类基因转染小鼠白血病细胞系(L细胞系)。这些受I类基因转染的细胞表面有I类抗原的表达。特异性Tc不能杀伤未经I类基因转染的病毒感染L细胞,但可杀伤经I类基因转染的病毒感染L细胞,表明转染后所表达的I类抗原起约束作用。Ozato等切割I类基因的不同片断进行基因杂交,并将其转染到L细胞系,成功地得到嵌合I类抗原分子,如α1及α2结构域取自与杀伤细胞I类基因相同的基因片段,而α3、穿膜及胞浆区取自与杀伤细胞I类基因不同基因片段,则杀伤细胞可杀伤病毒感染的靶细胞,所之则不然。上述试验证明,α1及α2结构域是参与杀伤和约束的有效部位。
利用突变品系小鼠研究发现靶细胞I类抗原突变分子的微小变化便能改变Tc细胞对它的识别。使如H-2kbml突变株(b为单倍体,m为突变)来自H-2Kb突变品系,只有个别氨基酸的差异,Kb约束病毒抗原特异性的CTL就不能识别和裂解多种病毒感染的Kb突变的bm1靶细胞,表明1~3个氨基酸的变化足以使I类抗原失去原有的约束作用(表6-16)。
3.MHC对Tc杀伤病毒感染靶细胞约束的机制 目前关于MHC对Tc细胞杀伤病毒感染靶细胞约束的机制一般认为是通过联合识别(associative recognition),即Tc表面一个受体识别MHC编码的抗原与病毒抗原的复合物。
Benacerraf认为MHC I类Ⅱ类抗耕牛具有双重功能,在诱导阶段通过其结合部位选择抗原决定基,激活Th、Ts、Tc等效应细胞;在效应阶段,靶细胞上的I类和Ⅱ类抗原又约束Th、Ts、Tc发挥效应。
(三)载体效应(cerrier effect)
Benacerraf认为载体不是单纯起运载半抗原的作用,还具有载体特异性。载体处于耐受状态的动物,虽给予半抗原载体复合物,也不能诱导产生抗半抗原的抗体。对半抗原再次应答的发生有赖于对半原记忆的B细胞和对载体记忆T细胞同时存在时才能发生(图6-19)。
表6-16 K(b)-约束的CTL(a)对Kb(b)突靶细胞上抗原的识别
CTL针对病毒抗原对Kb突变靶细胞的裂解
bm5(1)(c)bm6(2)bm9(3)bm3(2)bm8(3)bm11(1)bm1(3)
LCM病毒++++ +++ -
小鼠脱脚病病毒±+ + -
SV40++ ++++-
Moloney氏病毒++++++++-++-
Seudai病毒++++++++--
痘苗病毒++ ++ -
VSV++ (+)+ -
甲型流感病毒++ (+)- -
注:(a)CTL是Tc细胞,取自各种病毒致敏的小鼠脾,其本身的H-2型别为Kb;
(b)所有品系自B6突变而来,b为单倍型,m为突变;
(c)突变后氨基酸结构改变的数目
图6-19 载体效应
注:①第二次注射与第一次相同的半抗原(DNP)和载体(牛血清白蛋白BSA) ,结果产生对DNP的再次反应。
②第二次反注射的抗原中,半抗原与第一次相同,载体(卵白蛋白ovalbumin)与第一次注射抗原的载体不同,结果不产生再次反应;
③第一次注射后,单独注射卵白蛋白,经过一定时间后再注射载体为卵白蛋白、半抗原为DNP的抗原,结果产生再次反应。
Mitchison等载体效应的过继转移(adoptive transfer of carrier effect)试验,证实了上述关于载体效应的结论(图6-20)。
图6-20 载体效应的过继转移试验(Mitchison)
Raff等载体效应阻断实验进一步证实T细胞是载体特异的载体反应细胞(carrier-reactive cell),B细胞是半抗原反应细胞(hapten-reactive cell)(图6-21)。
图6-21 半抗原、载体反应淋巴细胞的鉴定(Raff等)
相关热词搜索:
上一篇:HLA的临床应用
下一篇:MHC参与免疫细胞识别抗原










频道总排行
医学推广

频道本月排行
- 247Ⅱ型补体受体(CR2)
- 90干扰素(IFN)
- 52Ⅲ型补体受体(CR3)
- 29B因子(factor B,Bf)
- 27集落刺激因子(CSF)
- 22粘附分子的种类和结构
- 18细胞因子的结构和生物学特征
- 17免疫球蛋白的多样性
- 14蛋白酪氨酸激酶
- 12I因子
热门购物
评论排行
- 2011年临床执业医师考试实践技能真...(13)
- 腋臭手术视频(12)
- 2008年考研英语真题及参考答案(5)
- 节食挑食最伤女人的免疫系统(5)
- 核辐射的定义和单位(5)
- CKD患者Tm与IMT相关(5)
- 齐鲁医院普外科开展“喉返神经监护...(5)
- windows7激活工具WIN7 Activation v1.7(5)
- 正常微循环(5)
- 美大学性教育课来真的 男女上阵亲...(4)
