- lncRNA���طΰ�ϸ��KRAS�źŴ��ݵ����ڻ���
- PLOS Medicine��ĸ����֢��ʳ�̶�����������ֵĹ�ϵ
- �ذ�����2.7%���人��Ⱥ������Ч�¹ڲ�������
- �ΰ�ɸ������Χ
- �����������?
- Ϊɶθ��Խ��Խ������������?
- ���Ѹ�֢��ʷ��Ůʿ������Ѫѹ
- ������������MTHFD1�����Ƽ�carolacton����Ч�����¹ڲ�������
- �ȡ�������ϸ������������������������
- �������һ�߽��ǣ�һ�ߺúû���ȥ

�ǵķֽ��л
2012-09-09 19:34:45 ��Դ��37��ҽѧ�� ���ߣ� ���ۣ�0 �����
(��)�ǽͽ�ĵ���
�����������������£������ڵĸ��ִ�л�ܵ��ϸ����ȷ�ĵ��ڣ�������������Ҫ�������ڻ������ȶ������ֿ�����Ҫ ��ͨ������ø�Ļ�����ʵ�ֵġ���һ����л�����������������淴Ӧ��ø���ƴ�л��Ӧ�ٶȣ�����ø��Ϊ����ø���ǽͽ�;������Ҫ����ø�Ǽ��Ǽ�ø(HK)�� ������Ǽ�ø-1(PFK-1)�ͱ�ͪ�ἤø(PK)��
����1.�����ĵ���
�����ȵ������յ����������Ǽ�ø��������Ǽ�ø����ͪ�ἤø�ĺϳɣ�����ٽ���Щø�Ļ��ԣ�һ����˵�����ִٽ����ñȶ�����ø�ı乹�����ε������������ñȽϳ־á�
����2.��л�������ø�ı乹����
����������������ø�У���������õ��Ǵ�Ч����͵�øPFK-1���������һ���ľ�����ʽ���������жԷ�Ӧ���� 6-������Ǻ�ATP�Ľ�ϲ�λ���������м������λ����������Ƽ���ϵIJ�λ��6-������ǡ�1��6��������ǡ�ADP��AMP���伤�������ATP�� ��������������Ƽ���ATP�ȿ���Ϊ��Ӧ�����ֿ���Ϊ���Ƽ�����ԭ�����ڣ���øһ��������Ϊ�����ATP���λ�㣬��һ��������Ϊ���Ƽ���ATP���λ �㣬����λ���ATP��������ͬ��������λ�������ߣ����Ƽ����õ�λ�������͡���ATP�����ֽ��λ�㣬��������ϸ����ATP����ʱ��ATP��Ҫ ��Ϊ��Ӧ�����֤ø�ٷ�Ӧ���У�����ϸ����ATP����ʱ��ATP��Ϊ���Ƽ���������ø��6-������ǵ�������
������������Ҳ����6-����������ữ���ɣ������ữ����C2λ������C4λ�������øҲ����һ����ø��������Ǽ�ø-2(PFK-2)��
����2��6����������ǿɱ��������������ø-2ȥ���������6-������ǣ�ʧȥ��������á�2��6����������ǵ�����������ǿ������Ǽ�ø-1��6-������ǵ�������ȡ��ATP����������(ͼ4��3)��

ͼ4-3 �ȵ���Ũ�����߶Ը�ϸ����2��6-���������Ũ�ȵ�Ӱ��
�����ٴ��ϱ�ͪ�ἤø�쳣���ɵ��������ǽͽ��ϰ�����ϸ���ƻ�������Ѫ��ƶѪ��
���������ǵ���������
���������������������£������ֽ����ɶ�����̼��ˮ�Ĺ��̳�Ϊ�ǵ���������(aerobicoxidation)�������������Ƿֽ��л����Ҫ��ʽ���������֯�е������Ǿ��������������ֽ������������
����(һ)������������
�����ǵ����������������ν��С���һ���������������ɵı�ͪ�ᣬ��ϸ��Һ�н��С��ڶ��������������в����� NADH+H+�ͱ�ͪ��������״̬�£������������У���ͪ������������������CoA����������ѭ����������������CO2��H2O��ͬʱNADH+H+�ȿ� �����������ݣ������������ữ��������H2O��ATP��������Ҫ�����������������������н��еĵڶ��δ�л��
����1.��ͪ�����������

�������������ȵ�ø�DZ�ͪ������øϵ(pyruvate dehydrogenase system)���˶�ø����������ͪ������ø����ø��TPP����������������ת��ø����ø�Ƕ����������øA�����ж�������������ø����ø��FAD���� �������������Һ�е�NAD+����ø�������γ��˽���������������Ӧ����������˴�Ч�ʡ�
�����ӱ�ͪ�ᵽ����CoA�������������йؼ��IJ����淴Ӧ���������Ӧ�ı�ͪ������øϵ�ܵ��ܶ����ص�Ӱ�죬�� Ӧ�еIJ������CoA��NADH++H+���Էֱ�����øϵ�еĶ�������������ת��ø�Ͷ�������������ø�Ļ��ԣ���ͪ������ø(pyruvate decarboxylase,PDC)������ADP���ȵ��صļ����ATP�����ơ�
������ͪ�����ⷴӦ����Ҫ�����DZ�ͪ�������ͷŵ�����������������CoA�еĸ����������У�������NADH+H+(ͼ4-4)��

ͼ4-4 ��ͪ������ø����������û���

����2.������ѭ��(tricarboxylic acid cycle)
��������CoA������һ������Ӧ���ɵ�ѭ����ϵ������������H2O��CO2���������ѭ����Ӧ��ʼ������CoA�� ��������(oxaloacetate)�������ɵĺ��������Ȼ��������ᣬ��˳�֮Ϊ������ѭ����������ѭ��(citric acid cycle)������ϸ�������£�
����(1)����CoA����������ѭ��
��������CoA���������������������㹻���������������Ȼ�����ȩ�������ϡ����ȴ�CH3CO���ϳ�ȥһ��H+�����ɵ������ӶԲ���������ʻ�̼�����˹���������������CoA�м��壬Ȼ�����������ˮ��ų�����������ᣬʹ��Ӧ����������ҽ��С��÷�Ӧ��������ϳ�ø(citrate synthetase)�����Ǻ�ǿ�ķ��ܷ�Ӧ��

�����ɲ������������CoA�ϳ���������������ѭ������Ҫ���ڵ㣬������ϳ�ø��һ���乹ø��ATP��������ϳ�ø�ı乹���Ƽ������⣬α-ͪ����ᡢNADH�ܱ乹��������ԣ�����֬��CoAҲ���������Ļ��ԣ�AMP�ɶԿ�ATP�����ƶ������á�
����(2)���������γ�
������������崼������������ת������������ʹ�崼����ٴ����������������˷�Ӧ��˳��ͷ��ø����Ϊһ���淴Ӧ��

����(3)��һ����������
������������������ø�����£�����������ٴ��������ʻ������ɲ���������(oxalosuccinate)���м� ���������ͬһø���棬������������α-ͪ�����(α��ketoglutarate)��NADH��CO2���˷�ӦΪβ-�������ȣ���ø��ҪMn2+��Ϊ �������

�����˷�Ӧ�Dz�����ģ���������ѭ���е����ٲ��裬ADP��������������ø�ļ��������ATP��NADH�Ǵ�ø�����Ƽ���
����(4)�ڶ�����������
������α-ͪ���������øϵ�����£�α-ͪ�����������������������CoA��NADH+H+��CO2����Ӧ������ȫ�����ڱ�ͪ������øϵ�����������ȣ�����α���������ȣ�����������������һ���ִ�����������CoA�ĸ����������С�
����α-ͪ���������øϵҲ������ø(α-ͪ���������ø����������������ת��ø����������������ø)�������ø(TPP�������ᡢHSCoA��NAD+��FAD)��ɡ�

�����˷�ӦҲ�Dz�����ġ�α-ͪ���������ø��������ATP��GTP��NAPH��������CoA���ƣ����䲻�����ữ/ȥ���ữ�ĵ��ء�
����(5)�������ữ����ATP
��������������ø(succinate thiokinase)�������£�������CoA��������ˮ�⣬�ͷŵ����������ںϳ�GTP����ϸ���ߵ������ֱ������ATP���ڲ��鶯���У�������GTP��������ATP����ʱ��������CoA�����������øA��
����
����(6)����������
��������������ø(succinate dehydrogenase)��������������Ϊ�Ӻ����ᡣ��ø�������������Ĥ�ϣ�������������ѭ����ø���Ǵ�������������еģ���ø�����������ĺ� ���۽�ϵ�FAD������������ĵ���ͨ��FAD���������ģ�Ȼ�������Ӵ�������O2����������������������������������øǿ�����ľ������������ �Կ������������ѭ����

����(7)�Ӻ������ˮ��
�����Ӻ�����ø�����Ӻ�����ķ�ʽ˫�������ã�����˳��ϩ����(������)�������ã�����Ǹ߶����������Եġ�

����(8)������������
������ƻ��������ø(malic dehydrogenase)�����£�ƻ�����ٴ��������������ʻ������ɲ�������(oxalocetate)��NAD+������ø�ĸ�ø���������ΪNADH+H+(ͼ4-5)��
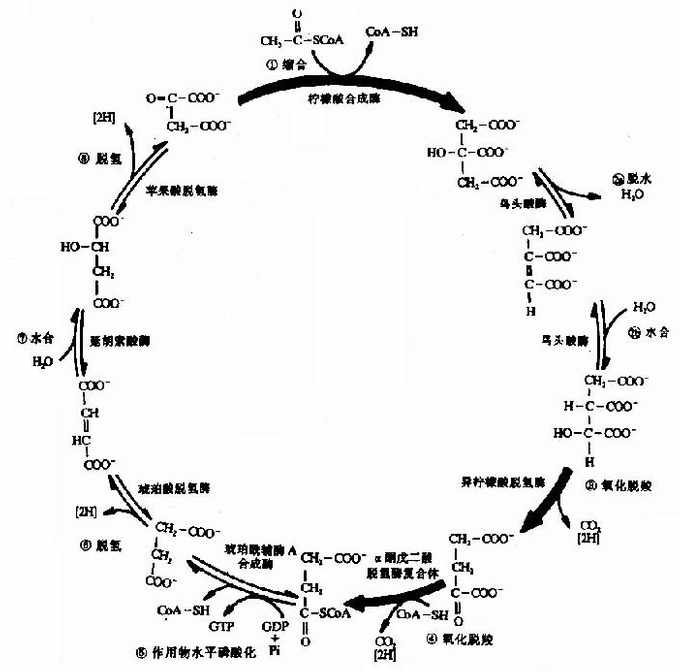
ͼ4-5��������ѭ��

����������ѭ���ܽ
��������CoA+3NADH++FAD+GDP+Pi+2H2O��—→
����2CO2+3NADH+FADH2+GTP+3H+ +CoASH
������CO2�����ɣ�ѭ�������������Ȼ���Ӧ(��Ӧ3�ͷ�Ӧ4)���ζ�ͬʱ���������ã������õĻ�����ͬ����������������ø������β���������ȣ���ø��NAD+��������ʹ�����������ɲ��������ᣬȻ����Mn2+��Mg2+��Эͬ�£���ȥ�Ȼ�������α-ͪ����ᡣ
����α-ͪ���������øϵ������α���������ȷ�Ӧ��ǰ����ͪ������øϵ���߾��ķ�Ӧ������ͬ��
����Ӧ��ָ����ͨ��������������CO2���ǻ����ڲ���CO2���ձ���ɣ��ɴ˿ɼ�������CO2������������ȼ������CO2�Ĺ��̽�Ȼ��ͬ��
������������ѭ�����Ĵ����⣬����������ԭ����NAD+Ϊ�����壬һ����FADΪ�����壬�ֱ�ԭ���� NADH+H+��FADH2�������־��������ڵ�����ϵ���ݣ����������������ˮ���ڴ˹������ͷų���������ʹADP��Pi�������ATP���� NADH+H+����ĵ�����ϵ��ÿ2H������һ����H2O������3����ATP����FADH2����ĵ�����ϵ������2����ATP���ټ���������ѭ������һ�� �������ữ����һ����ATP����ô��һ����CH2CO��SCoA����������ѭ����ֱ��ѭ����ĩ������12����ATP��
����������CoA����������̼ԭ�ӣ�����CoA����ѭ��������̼������Ӳ����������ϣ�������̼�������ᣬ��������ѭ�����ж�����������2����CO2�������ѭ���Ķ�̼��������̼ԭ������ȣ����ǣ���CO2��ʽʧȥ��̼��������������������̼ԭ�ӣ��������Բ������ᡣ
������������ѭ�����м����������Ͻ�������ѭ�������ģ���������ѭ���е�ijЩ��ɳɷֻ��ɲ���ϳ��������ʣ�����������Ҳ�ɲ���ͨ������;���������м�������˵������ѭ����ɳɷִ��ڲ��ϸ���֮�С�
�������硡�ݳ�������——→���Ŷ�����
����α��ͪ�����——→�Ȱ���
������������——→��ͪ��——→������
�������б�ͪ���Ȼ�ø�������ɲ�������ķ�Ӧ��Ϊ��Ҫ��
����
������Ϊ��������ĺ������٣�ֱ��Ӱ��ѭ�����ٶȣ���˲��ϲ������������ʹ������ѭ������˳�����еĹؼ���
����������ѭ�������ɵ�ƻ����Ͳ�������Ҳ�����������ɱ�ͪ�ᣬ�ٲ���ϳ������������ʻ��һ������(ͼ4-6)��

����(��)��������������������
����1.������ѭ���ǻ����ȡ��������Ҫ��ʽ��1�����������Ǿ������ͽ��������2������ATP�������������ɾ� ����38��ATP(���4?)������������ѭ������24��ATP����һ�����������£�������֯ϸ���Դ��ǵ�������������������ǵ�����������������Ч�� �ߣ����������ܣ���������ATP�����У�����ܵ�������Ҳ�ܸߡ�
����2.������ѭ�����ǣ�֬����������������Ҫ�л��������ڳ��������Ĺ�ͬ��л;����������ѭ������ʼ��������ø A���������������ֽ�����Ҳ������֬���ĸ��͡�֬��������Ե����ʵ�ijЩ�������л�����������ѭ��ʵ������������Ҫ�л����������������ܵĹ�ͬͨ ·������������2/3���л�����ͨ��������ѭ�������ֽ�ġ�
����3.������ѭ��������������Ҫ�л��ﻥ���������������Ǻ��������ڴ�л������α-ͪ����ἰ����������� ����ѭ�����м�����Щ�м�������ת���ΪijЩ���������Щ�������ֿ�ͨ����ͬ;�����α-ͪ�����Ͳ������ᣬ�پ���������;�������ǻ�ת��ɸ� �ͣ����������ѭ��������������Ҫ���л���ֽ��л�����չ�ͬ;��������Ҳ�����ǻ�������������
����(��)�����������ĵ���
������������������������Ϊ�����Σ���һ���ǽͽ�;���ĵ������ǽͽⲿ����̽�ֹ���������Ҫ���۵ڶ��α��� ������������������CoA������������ѭ����һϵ�з�Ӧ�ĵ��ڡ���ͪ������ø�����塢������ϳ�ø��������������ø��α-ͪ���������ø����������һ���� ������ø��
������ͪ������ø�������ܱ�λ����Ҳ�ܻ�ѧ���ε��أ���ø�����������Ĵ�����ATP������CoA��NADH���������ƣ����ֱ�λ���ƿɱ�����֬��������ǿ��������������ѭ��������CoA���٣���AMP����øA��NAD+�ѻ���ø������ͱ�
��λ�����������λ���ڣ� �ڼ����ﻹ�еڶ���εĵ��ڣ���ø���Ļ�ѧ���Σ�PDH���������ǻ�������һ���ǻ����ض���һ��˿����л������ữ��ø���Ծ������ƣ������ữ���� �ͻָ������ữ-�����ữ����������������ἤø�����ᵰ������ø�ֱ���ģ�����ʵ����Ҳ�DZ�ͪ��ø���������ɣ���ǰ�������ĵ��ڵ��ף���ø��ATP ��λ�����ATP��ʱ��PDH�����ữ���������ATPŨ���½�����ø����Ҳ���ͣ�������ø��ȥPDH�����ᣬPDH�ֱ������ˡ�
������������ѭ����������ϳ�ø��������������ø��α-ͪ���������ø�ĵ��ڣ���Ҫͨ������ķ���������ʵ�ֵģ� ��������ѭ���ǻ�����ܵ���Ҫ��ʽ�����ATP/ADP��NADH/NAD+���ߵı�ֵ������Ҫ�����ATP/ADP��ֵ���ߣ�����������ϳ�ø������ ��ø����ø���ԣ���֮ATP/ADP��ֵ�½��ɼ�����������ø��NADH/NAD+��ֵ��������������ϳ�ø��α��ͪ���������ø���ԣ�������ATP /ADP��NADH/NAD+֮������һЩ��л�����ø�Ļ���Ҳ��Ӱ�죬������������������ϳ�ø���ԣ���������CoA����α-ͪ���������ø���ԡ��� ֮����֯�д�л�������ѭ����Ӧ���ٶȣ��Ա���ڻ���ATP��NADHŨ�ȣ���֤��������������(ͼ4-7)��

ͼ4-6��������ѭ���л�ԭ��ø��CO2������

ͼ4-7��������ѭ�������Ƽ��ͼ����
��










Ƶ��������
ҽѧ�ƹ�

